Adaptive Plasticity as Causal Inference
Please find more from Charlie Munford at TalkingOctopus.

Genetic sequences are vitally important for life, but why? Is there a coherent view of biology other than that of The Selfish Gene? A scattered group of thinkers may be suggesting another way forward. Lynn Margulis, Peter Godfrey-Smith, Michael Skinner, James Shapiro, Eva Jablonka, Marion Lamb, Michael Levin, Richard Watson, and Denis Noble have all suggested various deep revisions of the idea that randomly mutating “selfish” genes contain instructions for life. But the selfish-gene view, narrow as it may be, is at least a great explanation. Could there be a philosophically coherent alternative idea to replace the elegant concepts of the Modern Synthesis?
In the selfish-gene view, all information in biology flows upwards from the genes to the phenotype. It posits that new variation arises through random genetic mutation, inheritance occurs through DNA, and natural selection of genes is the sole cause of adaptation, although genetic drift and spatial sorting are also present. This explanation always had an Achilles heel. It presupposes self-organization rather than providing a mechanism for it. Nothing could have been naturally selected until it was first self-organizing and reproductive. It is therefore a circular argument to hold that self-organization and reproduction are entirely explained by the natural selection of DNA. Another, perhaps unknown, process is required.
Natural selection is inherently subtractive. The philosopher of science Karl Popper once wrote an undated essay entitled, “A World without Natural Selection but With Problem-Solving,” a piece that was only published in 2014 in a book of criticism written by Hans-Joachim Niemann. In this thought experiment he proposed that we imagine an hypothetical expanding natural world with unlimited resources where every organism ever born never perished but was still alive today. He correctly pointed out that in this fictional world there would still be evolution. This philosophical problem was not enough to stimulate a re-evaluation, but empirical results have now challenged this genes-first view.
Lately a group of evolutionary biologists interested in the exceptions to these rules have gathered their observations into a concept called the Extended Evolutionary Synthesis. Among the authors of this view are Kevin Laland, Tobias Uller, Marcus Feldman, Kim Sterelny, Gerd Müller, Armin Moczek, Eva Jablonka, and John Odling-Smee. They have observed that developmental and phenotypic variation is non-random, that phenotypic variation can be assimilated into the genome, and that heritable traits can be passed on through paths other than DNA. In their view, causation is reciprocal between genes and the phenotype. These true observations, however, do not offer a good alternative explanation for the accumulation of complexity.
The beauty of the selfish-gene theory is that it neatly proposes a blind search whereby random chance could be converted, through natural selection, into a heritable molecule that could then instruct a life form. This mechanism would elegantly explain how complex function can accumulate in living beings. “Blind” here does not mean entirely random, or obeying no physical laws; it simply means that changes are uncorrelated with gains in functional adaptation. Its tenets have been elaborated into many complex mathematical models that often match experimental reality. A self-organizing set of particles already contain a profound bias, though; they are biased toward remaining in organized circulation as a self rather than dissipating entropically as a non-self. If this feature is presupposed, it leaves a large gap in the explanation.
In order to be a complete explanation, a theory has to be true in every case. A complete theory of the roles of DNA in living systems must match every example of DNA in living systems. For the selfish-gene idea to be sufficient, random genetic or phenotypic mutation would have to be the only source of heritable variation and natural selection the only form of instruction. There could be no bias toward adaptation except bias produced by the genetic code. As Popper (following David Hume) argued, no amount of proof in favor can protect a theory from refutation. The only thing that matters is whether examples can be found that prove it wrong. In this case, the sufficiency of the selfish-gene idea has been refuted by examples. This beautifully simple mechanism is incomplete. Random mutation is not the only source of heritable variation and natural selection is not the only means of instruction. So the question is now open: Where is the blind search?
If the blind search is not at the level of the genotype, then the only real alternative is that it is at the level of the phenotype. This idea is so strange and counterintuitive that it has not been seriously considered yet. How would this work, technically? So far it’s a total mystery. But there are several reasons we might suppose that the coding regions of DNA sequence (genes) don’t carry instructions at all. Instead, they may be just lists of templates for proteins that can be produced by the cell, if the cell chooses to do so. They may be like tools in a toolbox. The instructions that control which tools to use, and when, could possibly come from the environment around the individual cells and around organisms. This envelope of surrounding influences may be what tells all organisms what to become and what goals to pursue.
DNA is a part of the phenotype, and DNA sequences can turn off and on other sequences. So in these senses DNA could be said to be instructional. But every time you point to a DNA sequence as a cause of some phenomenon you have to ask the question, what activated this gene? The ultimate answer will always lie outside the genome, because the genome is only a part of the living system. I use the word “instruction” to mean the overall controlling logic of the system, not the action of particular instruments. A hammer instructs a nail to enter a board, in a sense, but something with knowledge of the entire building project instructs the hammer when and where to do so.
I believe it would strengthen the EES position to assert that organisms are shaped ultimately by downward causation, not reciprocal causation. Of course manipulating genes does cause phenotypic changes, so there is an immediate causal reciprocity in their role in the organism. But if genes are simply tools of a process whose function is ultimately instructed by the environment then the origin of complexity is in the interaction with downward causes at the level of the phenotype. Genes could merely be sophisticated instruments employed by the cell in its pursuit of phenotype-level goals. Genes cause functions only as parts of a functional phenotype.
The trouble with the EES position as it is now is that it is unclear where a blind search could occur. If genetic change is not random but biased toward adaptation, then this bias must be explained at a higher level. It makes little sense that the environment itself would contain a bias for life over non-life. Rather something about the organismal boundary at the phenotype level filters order from disorder in a way that produces that bias. Once you have broken the blind search at the level of the genome, you have to re-establish it somewhere else in order to preserve an atheistic view of the evolutionary process. Otherwise, why would a complex system store functional configurations?
To explain complexity, functional instructions have to pass through a filter and accumulate somewhere. If they are not accumulating in the genome, they have to be accumulating in the organism as a whole. The complexity of those instructions have increased over evolutionary time. Phenotypes, unlike genotypes, do not near-perfectly replicate themselves, but vary in form quite a bit based on the environment. If the instructions are contained not in the genotype but in the phenotype, they must be, on average, enhanced rather than degraded by phenotypic variation during active life. In order for function to increase in phenotypes during active life, there must be another filter in addition to natural selection.
Some, like Denis Noble and Eva Jablonka, have proposed that use and disuse is part of the process of inheritance. This has been termed Neo-Lamarckism, in honor of the eighteenth-century French biologist who assumed that use and disuse caused a heritable form of variation. This was also what Darwin himself and most other Victorian naturalists assumed. It is hard to deny that use and disuse do result in phenotypic changes. I suspect that this is the reason that higher animals require sleep as a repair process. They sleep to repair and reinforce the used parts and allow mutation in the disused parts in their organic beings. By “parts” I really mean processes, since an organism is really a set of nested, mostly circular processes.
The phenotype is a store of functional knowledge. Biases already present in the phenotype reduce the possible number of changes the organism can make in response to its environment, thereby shaping the evolutionary process. This can include developmental bias, niche construction, developmental plasticity, and behavioral adaptation. So much has been accurately pointed out by EES authors. But in order to account for increasing complexity we have to explain much more than this. We have to explain how new biases in favor of function are added to those already present. The only way this can happen is if there is a “blind” selection process at the level of the phenotype. By “blind” I do not mean that the phenotype is unaffected by its environment, quite the reverse. I mean that alterations in bias are not correlated with function until they are filtered by interactions with the environment. Although the phenotype has accumulated biases toward certain types of change, we are not concerned with those. We are concerned with the origin of changes to those biases.
Because of the EES, the phenotype level has a new, special burden to carry. The phenotype “knows” what instructions it contains already. But it doesn’t know in advance which aspects of its instructions will be mutated during active life. By blindly mutating (and then differentially repairing) its existing processes, some aspects of the phenotype must be reinforced and others discarded or mutated. But these changes tend to increase adaptation. Some subset of the results of that mutating process are inherited through multiple pathways of inheritance. The strange, counterintuitive conclusion this suggests is that the process of life is a matter of a phenotype engaging in a special type of causal predictive matching with its environment. Life itself, in other words, is a form of cognition. I nicknamed this blind search “epistolution” as a combination of the words evolution and epistemology, the sources of knowledge. A common name for this might be intelligence.
When the idea of developmental biases was introduced into evolutionary theory, we were still in familiar territory. Even Neo-Darwinists would readily accept that phenotypes have inherited bias. But when we assert that the instructions that produce this bias are not strictly in the (largely invariant) genetic molecule but in the plastic phenotype, then we have added a great new burden to the idea. Since phenotypes are highly malleable, we then have to explain how these biases are not only preserved through active life and inherited without degradation, they are informed and adapted by the process of active life. With this we are introducing a new philosophy entirely. The more accurate name for these biases then becomes goals. Life becomes about the evolution of goals. Variation becomes about the variations in goals, selection about the selection of goals, and instruction about the instruction of goals. Goals are much harder to explain because they are so manifestly derived from the immediate circumstances. And yet it is these immediate circumstances that determine which proteins are expressed by which genes in which cells at which times.
It is worth noting that even though we cannot prove this by building a machine (or even a computer program) that anticipates its environment well enough to adapt itself and save a library of its adaptations in molecular form like DNA, we also cannot build self-replicating molecules that evolve from non-living into living forms. So either explanation, both the upward-caused selfish-gene theory and a downward-caused EES/epistolution theory would require an undiscovered mechanism. In order to know for sure that self-replicators could even exist in principle, we would need to build them from non-living materials from scratch, and we have not been able to do this.
The epistolution proposal is the idea that an organism itself, in its adaptive plasticity in response to the world, is embodying a form of causal inference. It presents life as a struggle to understand unique niches, and only secondarily to survive and reproduce in them. This admittedly improbable idea may provide a new way to interpret the formation of biological selves and cellular goal-seeking, and could shed light on the still-murky definition of intelligence.
Several philosophers believe that the current roadblock to building genuine “thinking” AI is our lack of a fundamental theory of biological intelligence. The problem they propose is that all versions of machine “intelligence” today involve the programmers setting a goal function which produces output by a form of deductive logic (though often modeled on the concept of induction) from a set of training data. Yet organisms do not reason this way. The writer Erik Larson, quoting the philosopher C.S. Pierce, calls human intelligence a function of “abductive reasoning.” David Deutsch calls it “creativity.” The computer scientist Judea Pearl calls it “the ladder of causation.” Organisms, at least human organisms, make an observation, B, and then they invent an explanation for it, A, from their contextual understanding of the world. When more data, C, comes along, it either confirms or refutes their explanation. This has recently been proposed as the basis of the dopamine function in mammalian brains. The trouble is that no purely logical process of any sort will get you from B or C back to A, because this leap is non-logical. It is neither induction nor deduction. In its higher forms it is purely imaginative. It is, as Pierce is said to have called it, “broken deduction.” Causal inference requires something we haven’t discovered.
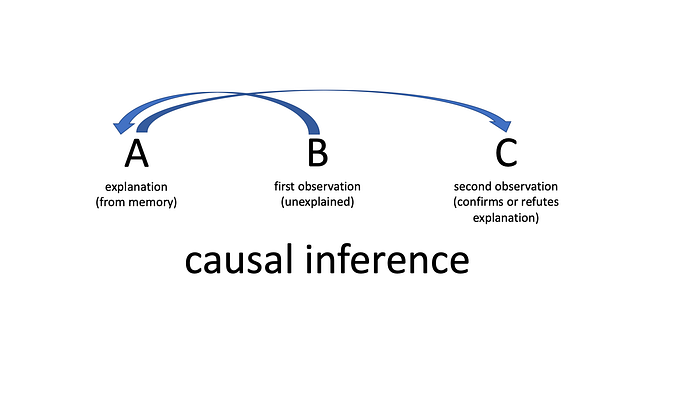
Today’s machine learning algorithms must all be programmed with specific goal functions, but the goals of cells and organisms shift about wildly depending on the context they find themselves in. A human stem cell may become a bit of bone, a neuron, a skin cell, and so forth depending on the context, just as a social mammal might pursue many different social or political roles within its group depending on the circumstances. The selfish-gene theory presented an organism as a machine with instructions and goals written into DNA that were aimed only at survival and reproduction. But this view has been refuted. Perhaps the goals of living beings are a direct outcome of their embodied explanations of the world. I call this “being is for knowing.”
…
My proposal is that the plasticity of an organism is an embodiment of causal inference, and this explains how plasticity can be adaptive. The basis of this idea is that genes are tools, and the instructions for how to use them come from the cellular or organismic environment. I came up with sixteen major reasons that this strange upside-down causation may be the right way to interpret cell biology:
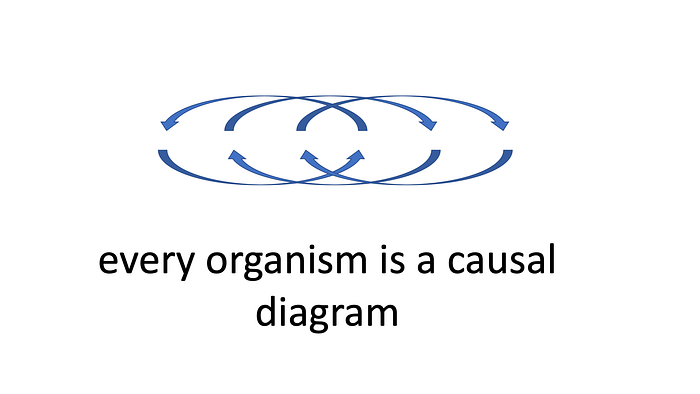
First, it is true that sustained reproduction and heredity depends, in all cases, on both cells and DNA. Even viruses replicate only by means of hijacking a cell’s gene expression mechanism. As Denis Noble points out in his book Dance to the Tune of Life, there are no conceivable biological experiments that can entirely separate these causal influences from one another. But he also suggests that there are examples of cells that can remain alive for a time without DNA, while DNA, without cells, is wholly inert. Red blood cells (erythrocytes) in the human body, for instance, are anuclear, containing no DNA at all. These cells are produced by a process of differentiation from erythroid stem cells in the bone marrow, and then live for 100–120 days in the bloodstream.
Second, we have discovered that all cells in our human bodies carry the same DNA even though there are 200 or more cell-types and they perform vastly different functions. Cells in large organisms like humans differentiate into myriad structures like bones, muscles, immune systems, neurons, and so on. This happens not because they carry different genetic sequences, but because different cellular environments during development cause different patterns of gene expression, making them diverge from a single pluripotent zygote into distinct tissues.
Third, we have discovered that cells in the same tissue express a given gene at markedly variable rates, with as much as 1000-fold difference in protein expression rates between two adjacent cells. Gene expression is stochastic at a small scale, and yet overall function is stable at a large scale. Organs function reliably, even while cells carry out their individual functions in highly variable ways. This result suggests that gene expression is determined by exterior conditions which are variable from cell to cell, rather than the invariant genetic molecule present in the nucleus of every cell. Most turtles and crocodilians depend on temperature for sex determination, indicating that exterior conditions can control development.
Fourth, we have discovered that the majority of genetic protein-coding sequences are likely to have no effect on phenotype by themselves under normal conditions. Gene knockout experiments often find extensive genetic buffering. In a study done by Hillenmeyer et al. in 2008 (The chemical genomic portrait of yeast) published in Science, over 80% of coding sequences silenced had no visible effect on the resulting organism. If the genome were instructional, this result would be puzzling. How could 80% of deleted vital instructions have no effect? If, however, the genome were a toolbox, then the possible redundancy and irrelevance to function of particular sections of DNA would be unproblematic. The role of informational templates in living systems could be “as-needed.” How much of the internet is useful? How many of the books in the Bodleian library are checked out on a regular basis?
Fifth, except in a few cases of rare genetic disorders, traits usually do not correspond to particular genes. Genome wide association studies that look for strong correlations between complex traits and specific genetic material usually fail to replicate. Without these strong correlations, how can selection target particular sequences to produce evolutionary change?The problem that traits correspond only very loosely with inherited genotypes is called the missing heritability problem. I can’t see how this problem is solved by the hypothesis that all genes are involved in complex traits. The Human Genome Project was hyped as a profound breakthrough for medicine, because it was predicted that knowing their genetic code would predict 80% or more of a human’s traits. But in truth, even the founders of these projects admitted some disappointment. Genetics is causally interrelated with all disease simply because all life requires proteins. These studies show that the genetic code does not always predict the way that genes are interpreted to produce phenotypes.
Sixth, experiments have been done to demonstrate that the germ cell can indeed have a large impact on development. For example, in a 2005 study of cross-species cloning, a common carp nucleus was transplanted into a goldfish recipient egg. The resulting fish had a vertebral column more closely resembling the cytoplasmic recipient than the nuclear donor. This demonstrates that the cell outside the nucleus can sometimes contribute significant instructions to development.
Seventh, we have discovered that transgenerational epigenetic inheritance (TEI) is found in many organisms. Nothing can be found until it is searched for, and investigations of this sort have only recently begun in earnest due to advancing molecular techniques. If the instructions were strictly in the genome, inherited epigenetic changes would degrade those instructions and quickly make them useless for maintaining life. It would be like a game of “telephone,” where intended messages quickly drift into nonsense. Even one genuine case of this would be sufficient to disprove the idea that the instructions for all life are contained exclusively in the genome. Yet examples of TEI have been found in complex model organisms across the tree of life: plants, yeast, worms, flies, even humans. In plants, for example, somatic cells can give rise to new germ cells, thereby sometimes passing along epigenetic changes they have acquired. In single-celled life it would be hard to imagine a way for TEI not to occur in nearly every lineage, since the entire parent cell structure is inherited by the daughter cells. There are complex, multicellular animals with nervous systems, planaria for instance, that reproduce from fission indefinitely. Without a proven step in all examples of inheritance that reprograms all germ cells by removing or randomizing all epigenetic marks, the integrity of the inherited message from generation to generation would disintegrate. These examples are plain disproof for the idea of genetic instructions.
Eighth, there is a clear mechanism for epigenetic changes to shape the form of the genome. This mechanism is known as “genetic assimilation,” and was discovered by Conrad Waddington in the 1950s. It was recently expounded by Mary Jane West-Eberhardt, who coined a phrase: “genes as followers.” Genetic assimilation happens when an organism first adopts a phenotype through epigenetic alterations, and then genetic change through natural selection reinforces or “canalizes” this change. By persisting in the lifestyle that the epigenetic changes permit, the organism causes the selection pressures that favor a genotype that supports the new phenotype. This mechanism allows the phenotype to change the genotype even without specifying particular molecular genetic sequences, by changing the selection pressure. The Central Dogma of molecular biology specifies that information only travels from DNA to protein and not back again. But Waddington’s mechanism transfers instruction to the genome without violating the prohibition against reverse translation from protein back to DNA.
Ninth, there is evidence that many organisms can take actions that directly reorder their genomes. Bacteria widely share genes outside their lineages, a phenomenon called “horizontal gene transfer.” Barbara McClintock discovered the phenomenon of “jumping genes” in maize in the 1950s, and further work has discovered other mechanisms that cells can sometimes use to directly change their gene sequences, especially when under stress. James Shapiro outlines many of these in his book Evolution: A View from the 21st Century. Our own immune cells even use a process of targeted hypermutation that helps produce matching antibodies for disease agents, although these changes are not directly heritable. But these examples show that phenotypes can directly instruct genotypes.
Tenth, there is a step in sexual reproduction called “crossing over” where the genetic material on each chromosome is cut and spliced independently to form a new combination of genetic material. This recombination does not respect any particular gene boundaries, ensuring that no sequence of genetic material can truly form an indivisible replicator. This means that sections of DNA are not, in themselves, units of Darwinian selection. The entire elegance of the argument for selfish genes is the idea of replicators getting themselves perfectly copied into the future as causes for complex phenotypic vehicles. But what, exactly, would the replicator be? The greatest strength of the genes-first idea was its specificity. The lack of a specific set of molecules that comprise the replicator in the selfish-gene theory is a conspicuous failure of the theory.
Eleventh, some organisms engage in niche construction, changing their environment to enable their lifestyles and reducing selection pressures that might force them to evolve different strategies. Beaver dams are a classic example, but there are many others: spider webs, coral reefs, prairie dog towns, termite mounds. These are ways that phenotypes act to instruct their own genes indirectly. As long as these had no molecular effect on genetic material this in itself is not a refutation of the genes-first idea. But if these interactions with the environment can become another pathway of inheritance, it does challenge the genes-first view.
Twelfth, organisms can also directly inherit functional behaviors from their parents outside of reproductive channels by imprinting and learning, as was demonstrated in a recent study of rat maternal licking. Jablonka and Lamb’s book Evolution in Four Dimensions is a great resource here. These downward causes are not accounted for in traditional evolutionary theory. The difficulty here is not that the behavior does not involve genes, of course it does involve genes. The difficulty is that the behavior itself is heritable, demonstrating that the genes do not contain instructions that control their own expression.
Thirteenth, we have discovered that the human gut microbiome is a self-organizing functional system akin to an organ. This microbial community is not inherited but largely acquired after birth. It forms in the human digestive tract out of foreign cells with foreign DNA. These communities change with diet, age, and other factors, yet the combination of bacterial taxonomic lineages remain as unique for each individual as a fingerprint. Despite thus containing totally divergent sets of nonhuman genetic material, these communities contribute to cognition, metabolism, immune function, and digestion. Health conditions causally attributed to imbalances in these communities can be every bit as serious as imbalances in somatic human cells. In this case, it is clear that a group of cells can be constrained into a functional community by their environmental cues without specific DNA that controls this form of organization directly.
Fourteenth, we now know that organisms that become less likely to die as they get older can have roughly the same number of genes as organisms that age rapidly. Take for example oak trees (genus Quercus) and octopuses (genus Octopus). The genomes of organisms in each genus contain roughly the same number of protein coding sequences, yet oaks can often live for hundreds of years while octopuses senesce in as little as one year. The best gene-centered explanation for senescence is the idea of antagonistic pleiotropy. This concept argues that since selection occurs on genes for beneficial effects on reproduction, genes that have a beneficial effect on young, reproductive organisms while causing deleterious effects on older organisms will be favored by natural selection. But this fails to explain why genes cannot, as in the case of the oak, favor reproductively successful older organisms, rather than, as in the octopus, degrading function quickly after reproduction. Some organisms, like certain jellyfish, age in reverse rather than senescing. These examples show that antagonistic pleiotropy could be an effect of lifespans rather than the cause. It’s possible that epigenetic changes that improve function, i.e. intelligent learning, explain the evolutionary fitness of rapidly senescing adaptive phenotypes in creatures like the Octopus genus.
Fifiteenth, we have discovered that phenotypes in the fossil record show not steady change based on random genetic mutation but an uneven rate of phenotypic change. If genetic sequences were the instructional basis for life, then we would expect that organisms would have changed relatively steadily as random mutations accumulated in their genes. Organisms have not changed steadily over geologic time; instead the fossil record shows long period of relative stasis interspersed by short periods of rapid evolution. There are bursts of creativity, such as the period known as the “Cambrian explosion” when the precursors to many of the major surviving groups of life forms first appeared. There is also recent evidence, such as in Darwin’s finches, that rapid evolution can occur in the present day. This evidence points to another, perhaps unknown mechanism, that underpins evolutionary processes beyond purely random genetic mutation.
Lastly, there is a phenomenon called “neutral molecular evolution.” This is when genetic change is not accompanied by phenotypic change. The fact that these two processes can be decoupled is clearly inconsistent with the genes-first view of evolution. A striking example was recently discovered in the DNA of the tuatara, the last remaining member of an order of reptiles called Sphenodontia whose fossils from the Cretaceous period closely resemble todays living lizards. To the researchers’ surprise, the rate of neutral molecular evolution in this species was the highest found in any vertebrate. This means that although the genetic sequences have been rapidly changing in the tuatara for perhaps 200 million years, the phenotype of the animal has remained relatively fixed. In this extraordinary case, the same sort of being has found new tools with which to do the same job. If genes really wagged the dog, so to speak, the tuatara would be a very different “dog” by now.
…
The question is not whether genes have a powerful effect on organisms. Everything in the cell is probably useful; otherwise it would not be there. The questions are what are the sources of variation? and what is the mechanism of filtering variation into functional forms? The universe can’t be biased in favor of life; there must be a mechanism to explain all adaptive bias. There is no doubt that natural selection filters organisms, but this single filter is not enough to account for adaptive plasticity resulting in increasing complexity. In this proposal the entire organism would be a storage unit for complexity, not only the genome. The genome would simply be the part most resistant to change. This information storage function is very helpful; we can imagine it as a tremendous library for complex templates of critical proteins. But a library does not tell a university what to read.
There is no doubt that at least one species of organism, Homo sapiens, is intelligent and creative. Biologists are also discovering much more about the adaptive learning capacities of other species. Crows, for example, can solve puzzles that five-year old children cannot solve. Dolphin and orca pods form distinct hunting cultures and communicate with songs that might resemble pre-human vocalizations. These capacities continue to make it harder to consider humans totally unique in our ability to learn and create knowledge in some form. Learning of any sort constitutes non-random change, instructed by the environment. Evolution is not necessarily aimed at complexity; organisms can shed complex traits as well as acquire them. But there is an acquired bias toward adaptation in phenotypes that then biases the selection of genomes. Where does this acquired bias come from?
There is currently no consensus definition of intelligence or creativity, and no scientific agreement on how far it reaches into the tree of life. Defining biological intelligence as “the ability to generate causal inferences” distinguishes it from known forms of computation, and makes it a testable, if not always quantifiable, criterion. Since we do not have a plausible theory for how this form of intelligence arises from groups of cells and we cannot build an intelligent being from non-living materials, we cannot rule out the possibility that causal inference is present in all life forms to some degree. If so, then it may be this ability that results in functional gene expression. Adaptive plasticity of any sort may be a form of causal inference about what the nature of the challenge to survival is and how an adaptive response should be mounted.
This idea entails an adaptive process aimed at survival and reproduction, but it adds a deeper layer of teleology. Survival and reproduction would be made possible by the fact that an organism is a process that seeks causal prediction. It presents life as a search for knowledge. Given what I am, what should I become, such that I can know? This would explain why humans can make causal inferences; it would be a property of all biological life to some degree. Our niche is complex, therefore our causal inferences are complex. But they might not be different in kind from those of other species.
This mystery may be an inspiring provocation to re-examine the natural world, and ourselves, to work out this mechanism.
Copyright March 1, 2023 by Charles S. Munford
